|
Terminology
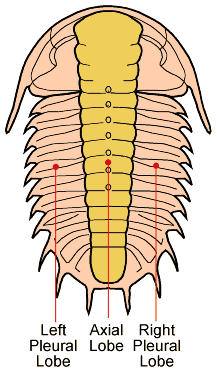 |
| "Trilobite" (meaning "three-lobed") named for the three
longitudinal lobes (Picture
Source) |
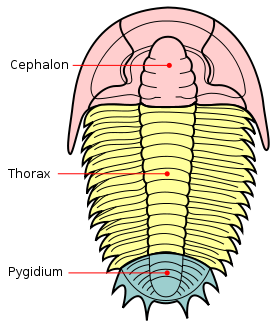 |
| The trilobite body is divided into three major sections (tegmata). |
As
might be expected for a group of animals comprising c.5,000 genera, the
morphology
and description of trilobites can be complex. However, despite morphological
complexity and an unclear position within higher classifications, there are
a number of characters that distinguish the trilobites from other
arthropods: a generally sub-elliptical,
dorsal,
chitinous exoskeleton
divided longitudinally into three distinct lobes (from which the group gets
its name); having a distinct, relatively large head shield
(cephalon) articulating axially with a
thorax comprising
articulated transverse segments, the hindmost of which are almost invariably
fused to form a tail shield (pygidium).
Physical Description
When trilobites are found, only the exoskeleton is
preserved (often in an incomplete state) in all but a handful of locations.
A few locations (Lagerstätten)
preserve identifiable soft body parts (legs, gills, musculature & digestive
tract) and enigmatic traces of other structures (e.g. fine details of eye
structure) as well as the exoskeleton.
Trilobites range in length from
1 mm to 72 cm (1/25 inch to 28 inches), with a typical size range of 3 to
10 cm (1 to 4 inches). The world's largest trilobite,
Isotelus rex, was found
in 1998 by Canadian scientists in Ordovician rocks on the shores of
Hudson Bay.
Exoskeleton
The
exoskeleton is
composed of
calcite and
calcium phosphate minerals in a protein lattice of
chitin that covers the
upper surface (dorsal) of the trilobite and curled round the lower edge to
produce a small fringe called the doublure. Three distinctive
tagmata
(sections) are present: cephalon (head);
thorax (body) and
pygidium (tail).
During
molting, the
exoskeleton generally split between the head and thorax, which is why so
many trilobite fossils are missing one or the other. In most groups facial
sutures on the cephalon helped facilitate molting. Similar to lobsters &
crabs, trilobites would have physically "grown" between the molt stage and
the hardening of the new exoskeleton.
Prosopon (surface sculpture)
Trilobite exoskeletons show a variety of
small-scale structures collectively called prosopon. Prosopon does not
include large scale extensions of the cuticle (e.g. hollow pleural spines)
but to finer scale features, such as ribbing, domes, pustules, pitting,
ridging and perforations. The exact purpose of the prosopon is not resolved
but suggestions include structural strengthening, sensory pits or hairs,
preventing predator attacks and maintaining aeration while enrolled.
In one example, alimentary ridge networks (easily visible in Cambrian
trilobites) might have been either digestive or respiratory tubes in the
cephalon and other regions.
Later, more advanced trilobites developed thicker cuticles (making
alimentary prosopon harder to see) against predation by cephalopods.
Spines
 |
| Ceratarges sp., an example of a species with elaborate spines
from the Devonian Hamar Laghdad Formation, Alnif, Morocco. |
Some trilobites such as those of the order
Lichida evolved elaborate
spiny forms, from the Ordovician until the end of the
Devonian period.
Examples of these specimens have been found in the
Hamar Laghdad Formation of
Alnif in Morocco.
There is, however, a serious counterfeiting and fakery problem with much of
the Moroccan material that is offered commercially. Spectacular spined
trilobites have also been found in western Russia; Oklahoma, USA; and
Ontario, Canada. These spiny forms could possibly have been a defensive
response to the evolutionary appearance of
fish.
Some trilobites had horns on their heads similar to those of
modern beetles. Based on the size, location, and shape of the horns the most
likely use of the horns was combat for mates, making the
Asaphida
family Raphiophoridae the
earliest exemplars of this behavior.
A conclusion likely to be applicable to other trilobites as well, such as in
the
Phacopid trilobite
genus
Walliserops
that developed spectacular tridents.
An
exceptionally well preserved trilobite from the
Burgess Shale. The
antennæ and legs are preserved as reflective carbon films.
An
exceptionally well preserved trilobite from
Beecher's
Trilobite Bed. Segmented legs are clearly visible. At this
Lagerstätte soft body
parts are preserved by
pyrite.
Digestive tract
The toothless mouth of trilobites was situated on the
rear edge of the hypostome (facing backwards), in front of the legs attached
to the cephalon. The mouth is linked by a small oesophagus to the stomach
that lay forward of the mouth, below the glabella. The "intestine" led
backwards from there to the pygidium.
The "feeding limbs" attached to the cephalon are thought to have fed food
into the mouth, possibly "slicing" the food on the hypostome and/or
gnathobases first. Alternative lifestyles are suggested, with the cephalic
legs used to disturb the sediment to make food available. A large glabella,
(implying a large stomach), coupled with an impendent hypostome has been
used as evidence of more complex food sources, i.e. possibly a carnivorous
lifestyle.
Sensory Organs
Many trilobites had complex eyes; they also had a pair of
antennae. Some
trilobites were blind, probably living too deep in the sea for light to
reach them. As such, they became secondarily blind in this branch of
trilobite evolution. Other trilobites (e.g.
Phacops rana and
Erbenochile erbeni)
had large eyes that were for use in more well lit, predator-filled waters.
Antennae
The pair of
antennae
suspected in most trilobites (and preserved in a few examples) were highly
flexible to allow them to be retracted when the trilobite was enrolled. The
antennae are
probably similar to those in extant
arthropods and as such
could have sensed touch, water motion, heat, vibration (sound), and
especially olfaction (smell) or gustation (taste).
Eyes
 |
|
The Schizochroal eye of
Erbenochile erbenii; the eye shade is unequivocal evidence that
some trilobites were diurnal.
|
Even the earliest trilobites had
complex, compound eyes with lenses made of calcite (a characteristic of all
trilobite eyes), confirming that the eyes of arthropods and probably other
animals could have developed before the Cambrian. Improving eyesight of both
predator and prey in marine environments has been suggested as one of the
evolutionary pressures
furthering an apparent rapid development of new life forms during what is
known as the Cambrian Explosion.
Trilobite eyes were typically
compound, with each lens being an elongated
prism. The number of lenses in such an eye varied: some trilobites had only
one, while some had thousands of lenses in a single eye. In compound eyes,
the lenses were typically arranged hexagonally. The fossil record of
trilobite eyes is complete enough that their evolution can be studied
through time, which compensates to some extent the lack of preservation of
soft internal parts.
Sensory pits
There are several types of prosopon
that have been suggested as sensory apparatus collecting chemical or
vibrational signals. The connection between large pitted fringes on the
cephalon of
Harpetida and
Trinucleoidea with
corresponding small or absent eyes makes for an interesting possibility of
the fringe as a "compound ear".
Fossil Record
The earliest trilobites known from
the
fossil record are
"Fallotaspids" (order
Redlichiida, suborder
Olenellina, superfamily
Fallotaspidoidea) and
Bigotinids (order
Ptychopariida, superfamily
Ellipsocephaloidea)
dated to some 520 to 540 million years ago. All trilobites are thought to have originated
in present day
Siberia, with
subsequent distribution and radiation from this location.
Trilobite fossils are found worldwide, with many
thousands of known species. Because they appeared quickly in geological
time, and moulted like other arthropods, trilobites serve as excellent
index fossils, enabling
geologists to date the age of the rocks in which they are found. They were
among the first fossils to attract widespread attention, and new species are
being discovered every year.
Origins
Early trilobites show all of the
features of the trilobite group as a whole; there do not seem to be any
transitional or ancestral forms showing or combining the features of
trilobites with other groups (e.g. early arthropods). Evidence suggests significant diversification
had already occurred prior to the preservation of trilobites in the fossil
record, easily allowing for the "sudden" appearance of diverse trilobite
groups with complex, derived characteristics (e.g. eyes).
Pre-Cambrian
Phylogenetic
biogeographic analysis of Early Cambrian
Olenellid and Redlichid trilobites suggests that a uniform trilobite fauna
existed over
Laurentia,
Gondwana and Siberia before the tectonic
breakup of the super-continent
Pannotia between 600 to 550 Ma. Tectonic
break up of Pannotia then allowed for the diversification and radiation
of the species. Break up of
Pannotia significantly pre-dates the first appearance of trilobites in the
fossil record, supporting a long and cryptic development of trilobites
extending perhaps as far back as 700 million years ago
or possibly further.
Paleozoic Era
Very shortly after trilobite fossils
appeared in the lower Cambrian, they rapidly diversified into the major
orders that typified the Cambrian -
Redlichiida,
Ptychopariida,
Agnostida and
Corynexochida. A Middle Cambrian and end
Cambrian extinction event marked major changes in trilobite fauna. During
the Ordovician trilobites were successful at exploiting new environments,
notably reefs. The end Ordovician extinction event again altered the
trilobite fauna, with 74% of the dominant Late Ordovician trilobite fauna
surviving into the Silurian. Drastic Middle and Late Devonian
extinctions almost wiped out the trilobites. Three orders and all but
five families were exterminated by the combination of sea level changes, and
other contributing factors, including a possible meteorite impact. Only a single order,
the
Proetida, survived into
the Carboniferous.
The Proetida survived for millions
of years, continued through the
Carboniferous period and lasted until the
end of the
Permian (where the vast
majority of species on Earth were wiped out). It is unknown why order
Proetida alone survived the Devonian. The Proetida maintained relatively
diverse faunas in deep water and shallow water, shelf environments
throughout the Carboniferous. For many millions of years the Proetida
existed untroubled in their
ecological niche. An
analogy would be today's
crinoids
which mostly exist as deep water species; in the Paleozoic era, vast
'forests' of crinoids lived in shallow near-shore environments.
Final Extinction
Exactly why the trilobites became
extinct is not clear, although it may be no coincidence that trilobite
numbers began to decrease with the appearance of the first
sharks
and other early
gnathostomes in the
Silurian and their subsequent rise in diversity during the Devonian period.
With repeated extinction events (often followed by apparent recovery)
throughout the trilobite fossil record, it is clear that more than one, or a
combination of causes is likely.
The closest extant
relatives of trilobites may be the
horseshoe crabs, or the
cephalocarids.
Trace Fossils
There are three main forms of trace fossils associated
with trilobites: Rusophycus; Cruziana & Diplichnites. These trace fossils
represent the preserved life activity of trilobites active upon the sea
floor.
Rusophycus, the resting
trace, are trilobite excavations which involve little or no forward movement
and ethological interpretations suggest resting, protection and hunting.
Cruziana, the feeding trace, are furrows
through the sediment, which are believed to represent the movement of
trilobites while deposit feeding. Many of the
Diplichnites fossils are believed to be
traces made by trilobites walking on the sediment surface. However, care
must be taken as similar trace fossils are recorded in freshwater and post
Paleozoic deposits, representing non-trilobite origins.
Gallery - From Wikipedia
|
|
|
Hypostome of
Isotelus sp., Ordovician age, southern Ohio, USA.
|
|
|
|
|
|
|
End of Reading

Return to the
Old Earth Ministries Online Earth
History Curriculum homepage.
Source:
Trilobite
|










{kind=link}
{kind=link}
{kind=link}
{kind=link}